1 ÚVOD
V této bakalářské práci se zaměřuji na popsání svalového vřeténka
a aplikaci teoretických poznatků do praxe v rehabilitačních metodách.
Bakalářskou práci rozděluji na část popisující svalové vřeténko a část
popisující užití jednotlivých rehabilitačních metod, které využívají funkce
svalového vřeténka.
V části popisující svalové vřeténko se zabývám teoretickými poznatky o struktuře, funkci, zapojení do neuromuskulárního systému a jeho zpětnovazebnou kontrolní funkci.
V části popisující užití jednotlivých rehabilitačních metod, které využívají funkci svalového vřeténka, se zabývám jejich popisem a využitím v praxi.
1. 1 Historické mezníky
Během minulého století byly informace o svalovém vřeténku
doplněny. Dle citace Hulligera závěry Sherringtona (1894) a Ruffiniho (1898)
vyřešily spor o tom zda jsou svalová vřeténka „svalovými pupeny“, které se podílejí
na tvorbě kosterních svalů nebo jsou-li dokonce ohniskem zánětlivých procesů.
Neočekávaným výsledkem, nyní však dobře známou skutečností bylo, že svalová
vřeténka jsou smyslové orgány, které jsou bohatě inervovány dostředivými i
odstředivými nervovými vlákny (Hulliger, 1983).
Ruffiniho klasické schéma ukazuje vřeténko jako útlý, prodloužený orgán sestávající se z malých, tzv. intrafusálních svalových vláken, která jsou v centrálním postavení uzavřena pouzdrem vytvořeným pojivovou tkání. Vydutí pouzdra dává orgánu typický vřetenovitý tvar a také jméno celému orgánu. Navíc, což je mnohem důležitější, obsahuje „smyslovou oblast” s hojným větvením a zakončeními dostředivých nervových vláken. Primární zakončení, jež jsou nyní nazývána Ia dostředivá vlákna, jsou ovinuta okolo intrafusálních vláken v místech procházejí centrální neboli „rovníkovou” oblastí. Sekundární zakončení (zakončení skupiny II dostředivých vláken) obsazují sousední juxta-equatoriální oblasti na obou stranách primárních zakončení (Hulliger, 1983; Trojan a kol., 1996).
Hulliger popisuje, že až za poměrně dlouhou dobu od prací Sherringtona (1894) a Ruffiniho (1898) byly
připsány různé funkční vlastnosti třem typům nervových zakončení vřeténka.
Objasnění fusimotorické kontroly svalového vřeténka začalo, dle
Hulligera, prací Leksella (1945) a Hunta & Kufflera (1951). Tito autoři
ukázali, že aferentní vlákna vřeténka
mohou být podrážděny selektivní stimulací gama-motoneuronů. Ty byly identifikovány díky jejich srovnatelně
pomalejší axonální přenosové rychlosti (15-55 m/s) na rozdíl od
alfa-motoneuronů (50-120 m/s).
V roce 1961 Cooper začal výzkum funkčních vlastností primárních a sekundárních receptorových zakončení. Klasifikoval funkčně jednotlivá aferentní vlákna vřeténka do skupin I a II na základě jejich axonální rychlosti přenosu. Jeho práce jasně demonstrovala, že skupina I byla mnohem více citlivá na rychlost svalového natažení než skupina II. Pozdější práce bohatě potvrdily tento rozdíl mezi dvěmi typy aferentních vláken v jejich rozdílné dynamické citlivosti na natažení svalu (Hulliger, 1983).
2 SVALOVÉ VŘETÉNKO
Složitá funkce uvědomování si polohy končetin jak vůči
ostatnímu tělu, tak i vzhledem k okolnímu prostředí, dále pak uvědomování
si svalového napětí, změny polohy, ale i rychlosti je zajišťována souborem
fyziologických mechanismů. Vlastní kontrola svalové funkce vyžaduje tedy nejen
excitaci svalu předními (anteriorními) motorickými neurony, ale také
nepřetržitou zpětnou vazbu mezi každým svalem a míchou přenášející senzorické
informace, které ukazují stav svalu v každém okamžiku pohybu. To znamená
např. jaká je délka svalu, jaké je konstantní napětí a jak rychle se délka a
napětí svalu mění (Hulliger, 1983; Trojan, 1996).
Aby mohly být tyto informace získány, jsou svaly a jejich
šlachy bohatě opatřeny dvěmi typy senzorických receptorů (Trojan a kol., 1996):
1.
Svalovými
vřeténky, jež jsou rozeseta na povrchu nebo uvnitř svalového bříška a posílají
informace do nervového systému, které popisují délku svalu nebo míru změny jeho
délky.
2.
Golgiho
šlachovými orgány, jež jsou umístěny ve svalových šlachách a přenášejí
informace, týkající se napětí šlachy či míry změny napětí.
Signály z obou těchto receptorů jsou buď úplně nebo téměř úplně vydávány za účelem vnitřní kontroly svalu. Jsou řízeny téměř vždy na podvědomé úrovni. Přenášejí ohromné množství informací nejen do spinální míchy, ale také do mozečku a dokonce i do mozkové kůry, a tak pomáhají každé této části nervového systému ve funkci kontroly svalové kontrakce.
Samozřejmě, že uvědomování si polohy končetin a i celého těla
prostřednictvím pouze těchto mechanismů, bez spolupráce ostatních smyslových orgánů (optického,
kožního a statoakustického analyzátoru) by bylo neúplné (Hunt, 1990).
2. 1 Struktura svalového vřeténka
Fysiologická organizace svalového vřeténka je ukázána na str.
12, obrázku 1 a příčný průřez svalovým vřeténkem na str. 15, obrázek 2. Každé
vřeténko je 3 až 10 mm dlouhé. Je
tvořeno okolo 3 až 12 velmi malými intrafusálními svalovými vlákny, jež jsou na
svých koncích zahrocena a připojena ke glykokalyxu větších extrafusálních
kosterních svalových vláken jež je obklopují.
Každé intrafusální svalové vlákno je velmi malým kosterním svalovým vláknem. Nicméně centrální oblast každého z těchto vláken – což je oblast uprostřed mezi oběmi konci – nemá buďto žádná, nebo jen velmi málo aktinových a myosinových vláken. Tento myotubulární segment vykazuje nahromadění jader a připomíná embryonální vlákno a na rozdíl od distálních částí není kontraktilní. Intrafusální vlákna jsou uložena v pouzdérku z pojivové tkáně. Pojivové pouzdro se ve střední části rozšiřuje a prostor mezi intrafusálními vlákny a pouzdrem je vyplněn lymfou. Oba polární konce intrafusálního vlákna jsou kontraktilní a mají charakter kosterního svalu. Tyto koncové části, které se stahují, jsou drážděny malými gama- motorickými nervovými vlákny, nacházejících se v předních rozích spinální míchy, jak již bylo popsáno dříve. Tato vlákna jsou také nazývána gama eferentní, na rozdíl od velkých alfa eferentních vláken (původem z alfa nervových vláken), jež inervují extrafusální kosterní svaly (Hunt, 1990; Matthews, 2000; Trojan a kol., 1996; Trojan, Druga, Pfeiffer & Votava, 2001).
Aferentní neurony začínají v centrální oblasti
vřeténka ze sensorických zakončení umístění na intrafusálních vláknech
uvnitř pouzdra. Tato centrální neboli senzorická oblast je dohodou rozdělena na
rovníkovou oblast – equatoriální (obsazenou primárními zakočeními velkých a
rychle přenášejících Ia aferentních vláken) a juxtaequatoriální oblast
(obsazenou sekundárními zakončeními menších a pomaleji přenášejících
aferentních vláken skupiny II) (Guyton & Hall, 2000; Deursen, Sanche,
Ulbrecht & Cavanagh, 1996).
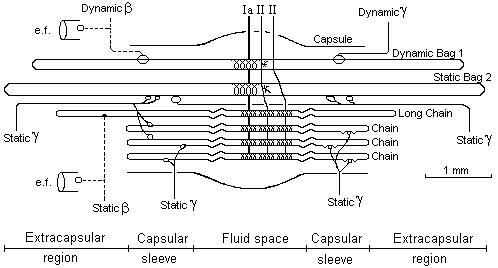
Obr. 1 Svalové vřeténko
Vysvětlivky
k obr. 1: Extracapsular region – mimokapsulární oblas
Capsular sleeve –
kapsulární obal
Fluid space –
tekutý prostor
Static –
statické vlákno
Dynanic –
dynamické vlákno
Popis obr. 1 :
Kapsulární obal
Prostřední
(equatoriální) část svalového vřeténka je přibližně stejně tak dlouhá jako
chain vlákna je někdy podobná hemencefalické bariéře. Udržuje proteiny a chrání
mechanicky senzorová zakončení. Je naplněna lumbrikační tekutinou.
Dynamická bag 1 vlákna
Dynamická bag
1 vlákna intrafusální jsou histologicky klasifikována jako bag1 vlákna (mají
více jak 100 jader v centrální části a méně ATPazy). Dynamická se nazývají
kvůli mechanickým vlastnostem (velice rychlá odpověď na změny délky svalu).
Statická „bag“ 2 vlákna
Statická „bag“
2 vlákna – intrafusální vlákna jsou histologicky klasifikována jako bag II.
vlákna (do 100 jader v centrální oblasti). Nazývají se statická, kvůli
jejich mechanickým vlastnostem (odpověď na změny svalové délky je pomalá).
Dlouhá „chain“ vlákna
Dlouhá „chain“
vlákna - obyčejně jedno „chain“ vlákno je delší než ostatní a je inervováno
statickým beta axonem.
„Chain“
vlákna
„Chain“ vlákna jsou kratší než „bag“ vlákna. Většina sekundárních zakončení (II aferents) přichází z „chain“ vláken. Fusimotorická stimulace gama-statických vláken je nejvíce efektivní mezi 50 – 150 Hz a sestává se z lokalizovaných kontrakcí, které přicházejí z obou primárních a sekundárních zakončení.
Ia aferent
Skupina Ia
aferentu přivádí informace z centrálního anulospirálního zakončení všech
intrafusálních vláken. Ia aferent má silný excitační efekt (mono nebo
polysynaptický) na alfa motoneuronech stejného svalu a jeho synergistech. Má
inhibiční efekt na svaly antagonistů.
II aferent
Skupina II
aferentních vláken přivádí senzorické informace ze sekundárních zakončení
většinou se sdružujících v „chain“ vláknech.
Statické gama-motoneurony
Statické
gama-motoneurony jsou tak nazývány kvůli svému výslednému efektu na
intrafusálních vláknech (statická „bag“ 2 vlákna a „chain“ vlákna). Když je
statický intrafusální systém aktivní, citlivost na změnu délky primárního
zakončení je redukována.
Dynamické gama-motoneurony
Dynamické gama-motoneurony jsou nazývány podle svého efektu na intrafusálních vláknech (dynamický „bag“ 1 vlákna). Když je dynamický intrafusální systém aktivní, citlivost na změny délky primárních zakončení je zvýšený.
„Chain“ vlákna
„Chain“ vlákna
– primární senzorické zakončení. „Chain“ vlákna mají centrální anulospirální
zakončení, tvořená Ia aferentními vlákny.
2. 2 Intrafusální vlákna
Intrafusální vlákna jsou svými úpony zapojena paralelně s vlákny extrafusálními. Intrafusální vlákna nejsou stejná. V tomtéž svalovém vřeténku nacházíme intrafusální vlákna lišící se délkou, průměrem, histochemickými vlastnostmi a uložením jader v centrálním myotubulárním segmentu. Rozlišit intrafusální svalová vlákna lze podle vlastností pod světelným či elektronovým mikroskopem a podle jejich mechanických a elektrických vlastností (Guyton & Hall, 2000; Trojan a kol., 1996).
Rozdělení intrafusálních vláken (Guyton & Hall, 2000;
Trojan a kol., 1996)
1.
Vlákna
(1 – 3 na vřeténko) silná, dlouhá, s vakovitým ztluštěním myotubulárního
segmentu, tzv. vlákna s vakem I. typu (nuclear bag1) a krátká, tenčí
s menším množstvím jader, tzv. vlákna s vakem II. typu (nuclear bag2)
přibližná velikost bag1 vláken je 20-25μm.
2.
Vlákna
(3 – 9 na vřeténko) tenká s řetězovitým uložením jader (nuclear
chain) o velikosti poloviční
10-12μm.
Tyto morfologické rozdíly itrafusálních vláken jsou jedním
z faktorů, které podmiňují
rozdílnou citlivost svalového vřeténka a umožňují diferencovat dynamická
a statické stimuly.
Pro všechny typy vláken platí, že hromadění jader je přímo
úměrné snížené hustotě kontraktilního materiálu (myofibril) (Hulliger, 1983).
Základním rozdílem mezi vlákny bag1 a bag2 je jejich
histochemické složení v části pro myosin ATP-asu.
Ve svalových vřeténkách existují tedy intrafusální svalová
vlákna tvořící více méně dva oddělené systémy. Jeden z nich je nastaven na
vnímání dynamických dějů (rychlost změny svalové délky) a je charakterizován
rychlou adaptabilitou, druhý na vnímání statických dějů.
Primární Ia nervové zakončení může být podrážděno jak nuclear
bag intrafusálními vlákny, tak i nuclear chain vlákny. Naopak sekundární II
zakončení je obvykle podrážděno pouze nuclear chain vlákny (Boyd, 1998; Trojan
a kol., 1996).
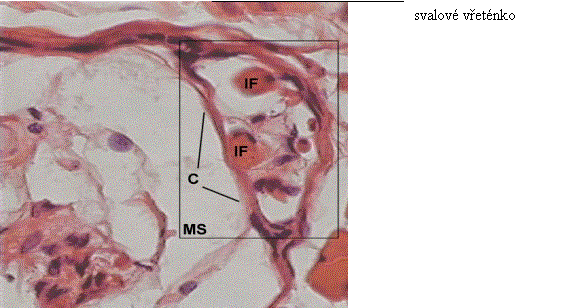
Obr. 2 Příčný průřez svalovým vřeténkem
2. 2. 1 Inervace
Intrafusální vlákna svalových vřetének mají trojí inervaci –
senzorickou, motorickou a autonomní.
2. 2. 1. 1 Senzorická inervace
Receptorická část svalového
vřeténka je umístěna v centrální
části, kde nemají intrafusální svalová
vlákna části kontraktilní. Jsou
stimulovány natažením této střední části
svalového vřeténka. A je lehce možné pozorovat, že mohou být podrážděny
dvěma způsoby (Chaitow, 1996; Guyton & Hall, 2000):
1)
Prodloužení
celého svalu natáhne střední část
intrafusálních vláken vřeténka, a tím podráždí
receptor.
2)
Dokonce,
i když nedojde ke změně celkové délky svalu, stažení koncových částí vřeténkových intrafusálních vláken také natáhne střední část těchto vláken, a tak opět dojde k podráždění
receptoru.
V centrální oblasti svalového vřeténka se nacházejí dva
typy senzorických zakončení. Jsou to primární a sekundární zakončení. Vychází
jednak z centrálních anulospirálních zakončení v primárním
myotubulárním segmentu, jednak z periferní části tzv. sekundární
myotubulární vlákna (keříčkovité zakončení). V samém středu receptorické
oblasti obkružuje centrální oblast
každého intrafusálního vlákna velké senzorické nervové vlákno, jež vytváří tzv.
primární neboli anulospirální zakončení. Některá senzorická vlákna mají svá
zakončení ve stěně pouzdra svalového vřeténka nebo pronikají až do pojivové
tkáně mezi intrafusálními vlákny. Toto nervové vlákno je typu Ia
s průměrem 10 – 17 μm, rychlost vedení vzruchu do spinální míchy je
mezi 70 – 120 m/s a je při výstupu z pouzdra opatřeno myelinovou pochvou.
Rychlost vedení vzruchu je obdobné jako pro jakýkoliv jiný typ nervového vlákna
v celém těle (Guyton & Hall, 2000; Hulliger, 1983; Matthews, 2000).
Receptorická oblast na jedné či obou stranách primárních zakončení je inervována obvykle jedním nebo dvěma menšími senzorickými vlákny typu II, o podstatně menším průměru 6 - 8 mikrometrů, jak je vidět na str. 18, obr. 3. Tato senzorická zakončení jsou nazývána sekundárními zakončeními; někdy okružují intrafusální vlákno stejným způsobem jako to dělají vlákna typu Ia, ale často se jen rozšiřují jako „větve na keři“ a jsou proto nazývána buď „trail“ zakončením (ocasovitým) nebo také „flower – spray“ zakončením (rozprašovacím) (Hulliger, 1983; Trojan a kol., 1996).
Vlákna ze sekundárních myotubulárních zakončeních jsou rovněž myelinizována a rychlost jejich vedení vzruchu je 30 – 70 m/s.
Svalová vřeténka nemusí být zásobena oběma typy senzorické
inervace. Při nepřítomnosti aferentních vláken vycházejících ze sekundární
myotubulární oblasti, mluvíme o svalovém vřeténku jednoduchého typu.
2. 2. 1. 2 Motorická inervace
Je realizována
fusimotorickými vlákny tzv. Gama (γ) systému a vlákny beta (β)
systému. Motorická vlákna vstupující do svalových vřetének patří k tzv.
Gama systému (fusimotorická vlákna) a končí v motorických ploténkách
kontraktilních segmentů intrafusálních bag1 vláken (3 – 6 μm, rychlost
vedení vzruchu 30 – 70 m/s), nebo vytváří difúzní vícečetná zakončení na
vláknech bag2 a na řetězových vláknech (Hulliger, 1983; Trojan a kol., 1996).
Vlákna beta-systému jsou
myelinizovaná vlákna stejného typu jako alfa-motoneurony, inervují současně jak
intrafusální, tak extrafusální svalová vlákna. Zatímco většina nervových
vláken, senzitivních i motorických, vstupuje do svalového vřeténka
v centrálním segmentu, beta-motorická vlákna vstupují do svalového
vřeténka v polárním segmentu a končí v motorických ploténkách bag1
vláken (Boyd, 1998).
Statická fusimotorická nervová vlákna typu gama inervují
intrafusální vlákna typu bag2. Příslušná aferentní senzorická vlákna
vycházející ze sekundárních myotubulárních zakončení patří k vláknům II,
pomalu vedoucí vzruchy.
Dynamická fusimotorická nervová vlákna gama a beta mají
zakončení v ploténkách intrafusálních svalových vláken bag2. Senzorická
vlákna vycházejí z primárních anulospirálních zakončení a jsou to velmi
rychle vedoucí vlákna Ia (Boyd, 1998; Hulliger, 1983).
2. 2. 1. 3 Autonomní inervace
Vedle senzorických a
motorických vláken nacházíme ve svalovém vřeténku i postgangliová sympatická
vlákna (0.3 – 1,3 μm, rychlost vedení vzruchů 0,7 – 2,3 m/s). (Hulliger,
1983)
![]()
![]()
Obr. 3 Schéma inervace svalového vřeténka
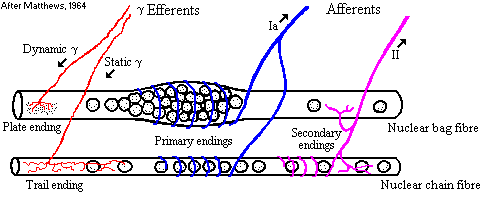
Vysvětlivky
k obrázku 3: afferents – aferentní vlákno
efferents – eferentní vlákno
dynamic
γ – dynamické gama vlákno
nuclear
bag fibre – vlákno nuclear bag
nuclear chain fibre –
vlákno nuclear chain
primary
endings- primární zakončení
secondary endings – sekundární
zakončení
static
γ – statické gama vlákno
plate
ending – plošné zakončení
trail ending –
„ocasovité“ zakončení
2. 3 Deformace jako podnět podráždění
Podnětem podráždění svalového vřeténka je deformace
myotubulárního segmentu intrafusálních vláken v důsledku protažení
extrafusálních svalových vláken. Práh dráždivosti svalových vřetének je velmi
nízký, dovedou zaregistrovat prodloužení svalových vláken o 0,05 mm (Guyton
& Hall, 2000; Opavský, 2002).
Deformace myotubulárního segmentu intrafusálních vláken je
bezprostřední příčinou podráždění senzorických zakončení, jejichž elektrickým
projevem je receptorový potenciál. Velikost receptorového potenciálu je přímo
úměrná stupni protažení extrafusálních vláken. Když receptorový potenciál
dosáhne „spouštěcí“ úrovně, vznikají v aferentních senzorických vláknech
akční potenciály, v nichž je zakódována informace získaná receptorem o
změně prostředí, v našem případě o protažení svalu (statická komponenta) a
jeho rychlosti (dynamická komponenta) (Hulliger, 1983; Hunt, 1990).
Velmi nízký práh dráždivosti svalových vřetének je významným
faktorem pro zajištění svalového tonu. Minimální změny délky svalových vláken
v době „svalového klidu“ jsou stálým zdrojem aktivace primárních i
sekundárních zakončení senzorických vláken. Aferentace z těchto receptorů
spolu s účastí gama-systému zajišťuje stálý klidový tonus svalových vláken
i po velmi dlouhou dobu (prostřednictvím gama-systému se projevuje tonizující
vliv CNS). U nižších obratlovců kontrolní funkci gama-systému plní
beta-neuronový systém, který neztratil svůj význam ani u savců, kde se účastní
na koaktivaci intrafusálních a extrafusálních vláken svalového vřeténka
(Hulliger, 1983; Trojan a kol., 1996).
Mezi faktory ovlivňující tzv. fyziologické zpoždění mezi
podrážděním a mechanickou odpovědí patří především: převaha typu vláken ve
složení svalu, rychlost pálení motorické jednotky, viskoelastické vlastnosti
svalu a šlachových tkání (Rodová, Mayer a Janura, 2001).
2. 4 Funkční vlastnosti vřeténkových a fusimotorických neuronů
Základní funkční vlastností
neuronu svalového vřeténka jsou (Hulliger, 1983):
1.
jako
doplněk ke své statické (polohové) citlivosti mají primární aferentní vlákna
(Ia) vysoce dynamickou citlivost na natažení svalu
2.
u
sekundárních aferentních vláken (II) je statická citlivost mnohem výraznější
než citlivost dynamická
3.
dynamické
fusimotorické neurony (beta a gama) dále zvyšují dynamickou citlivost
primárních aferentních vláken
4.
kromě
mohutného dráždivého účinku snižují statické fusimotorické neurony (γ)
dynamickou citlivost mnohem zřetelněji u primárních (Ia) než u sekundárních
(II) aferentních vláken
2. 5 Odezva receptorů na protažení svalového vřeténka
2. 5. 1 Dynamická citlivost: primární, sekundární a přechodní aferentní
vlákna
Aferentní vlákna svalového
vřeténka je snadné podráždit natažením mateřského svalu, jestliže je sval
natažen o konstantní délce jsou odpovědi primárních (Ia) a sekundárních (II)
aferentních vláken v podstatě stejné, kromě větší variability výbojů,
která je typická pro primární aferentní vlákna. Takže tyto dva typy aferentních
vláken se od sebe příliš neliší v polohové citlivosti. Funkčně lze hlavní
rozdíl mezi primárními a sekundárními aferentní vlákna nalézt v dynamické
citlivosti při natažení svalu. Tedy
během dynamické fáze napnutí a udržování napětí svalu reagují primární
aferentní vlákna nestále, což je mnohem nápadnější než u sekundárních
aferentních vláken. Obecně platí, že reakce primárních aferentních vláken (Ia)
jsou citlivější na polohu, rychlost a zrychlení, zatímco sekundární aferentní
vlákna (II) reagují jen na polohu a rychlost složek podnětu (Hulliger, 1983).
Mezi rychlostí axonálního přenosu a dynamickou reakcí nacházíme
značný rozptyl, což je částečně zapříčiněno existencí vřeténkových aferentních
vláken s reakčními vlastnostmi, ležícími mezi primárními a sekundárními
aferentními vlákny. Tato přechodná aferentní vlákna dosahují také často
středních hodnot rychlosti axonálního přenosu (okolo 72 m/s) (Hulliger, 1983;
Trojan a kol., 1996).
2. 5. 2 Citlivost svalových vřetének na pohyby
2. 5. 2. 1
Lineární vlastnosti svalových vřetének
Při používání metod, které by analyzovaly odpovědi vřetének na pohyb, se vycházelo z popisu lineárního přenosu funkcí. Tato očekávání nebyla úplně naplněna, protože odpovědi vřetének vykazují nápadnou nelinearitu. Při tzv. lineárním rozpětí má citlivost (poměr mezi reakcí a podmětem) primárních aferentních vláken tendenci zůstávat přibližně konstantní (Hulliger, 1983).
Nicméně za hranicí tohoto lineárního rozpětí citlivost
s dalším zvyšováním velikosti podnětu klesá. Takovéto nelineární chování
je typické jak pro primární, tak pro sekundární aferentní vlákna, ať ve stavu
pasivním nebo během aktivace statických nebo dynamických fusimotorických vláken
(Hulliger, 1983).
Hulliger (1983) uvádí, že při systémové analytické metodě, při
dráždění primárních aferentních vláken, se hodnoty lineárního rozpětí malých
pohybů pohybují v řádu 500 impulsů / s / mm. Takže citlivost při malých
pohybech může být až 100 krát vyšší než statická polohová citlivost. Pasivní
sekundární aferentní vlákna jsou asi 5 krát méně citlivější než primární
aferentní vlákna (100 impulsů /s /mm), nicméně statická fusimotorická akce tyto
rozdíly převážně smazává. Tyto poznatky platí pro rozpoznání citlivosti
primárních aferentních vláken na pohyby s velmi malou amplitudou.
Vřeténková aferentní vlákna a zvláště jejich primární jednotky jsou tedy vysoce
citlivými detektory malých pohybů, zatímco jejich reakce na velká natažení jsou
srovnatelně slabší (Doursen, 1996; Hulliger, 1983; Trojan a kol., 1996).
Vysoké hodnoty citlivosti na malé pohyby, které byly pravidelně
nacházeny při výzkumech, používající drobná natažení jež byla aplikována při
stálých délkách svalu, se jen velmi málo podobá přirozeným pohybům, které
většinou patří mimo úzký lineární rozsah. Při pohledu na chování vřeténka
v průběhu pohybů s velkými amplitudami se ukázalo, že citlivost na
malé pohyby může být drasticky snížena (na 5 – 20 % kontrolních hodnot při
konstantní svalové délce). Navíc bylo zjištěno, že rozsah tohoto snížení
citlivosti během pohybů závisí na rychlosti pohybů, a že vysoké hodnoty
citlivosti jež byly získání při konstantní průměrné délce byly dosahovány jen
při velmi pomalých pohybech. Tyto údaje naznačují, že domnělá role primárních
vřeténkových aferentních vláken jako zvláště citlivých detektorů malých poruch
během volních pohybu, může být spornější než se původně myslelo: pokud se sval
nehýbe velmi pomalu, není citlivost pasivních primárních aferentních vláken na
malé pohyby vyšší než pro pohyby mimo klasický lineární rozsah (Guyton &
Hall, 2000; Hulliger, 1983).
2. 5. 2. 2 Nelineární
vlastnosti svalového vřeténka
Nelinearita komplikuje
používání matematických modelů pro vyhodnocování role vřeténkových aferentních
vláken při kontrole pohybu. Při kvalitativní analýze zvyšuje prostor pro
dedukce vycházející z výbojů vřeténkových aferentních vláken, které
umožňují odhadnout fusimotorickou aktivitu během přirozených pohybů. Mezi
některé příklady nelineárního chování vřeténka můžeme zařadit např. závislost
na délce, rychlost odpovědi a okluze (Hulliger, 1983).
Závislost na délce - citlivost na malé pohyby závisí také na
průměrné délce svalu. Tato závislost může být tak výrazná, že citlivost u
ochablého svalu může být 10 krát menší než u svalu napnutého. Velikost tohoto
účinku je stejně velká jako snížení citlivosti vyvolané silnou statickou
fusimotorickou akcí nebo doprovodnými pohyby o velkých amplitudách. Rozsah
závislosti citlivosti k malým pohybům na délce je redukován jak statickými,
tak i dynamickými fusimotorickými účinky. Fusimotorické neurony mohou, tedy
pokud jsou vhodně aktivovány, kromě udržení palby vřeténka během zkracování
svalu, také udržet citlivost na malé pohyby na přibližně konstantní úrovni
(Boyd, 1998; Trojan a kol., 1996).
Okluze - v průběhu kombinované stimulace statických i
dynamických fusimotorických neuronů, může být statický účinek úplně zastíněn
účinkem vyvolaným samotnou dynamickou akcí (Hulliger, 1983).
Stimulace fusimotorických neuronů může zanechat vřeténko ve
změněném stavu. Tento stav může trvat až několik minut, pokud není odstraněn
velkými nataženími. Následkem může být zvednutí klidového výboje a jako nápadné
posílení úvodního výbuchu na začátku rychlého natažení nebo jako zvýšení
citlivosti Ia aferentních vláken na malé pohyby během pomalých doprovodných
pohybů o velké amplitudě (Boyd, 1998; Trojan a kol., 2001).
2. 6 Statické a dynamické fusimotorické neurony
Původní rozdělení
fusimotorickýn neuronů na statická a dynamická vlákna bylo založeno na spíše
funkčních než morfologických kritériích.
Toto dělení je stejné i na základě jejich potlačování či podporování
velikosti dynamické odpovědi Ia aferentních vláken na natažení aplikované na
mateřský sval. Prudké natažení a držení bylo často používáno pro roztřídění
fusimotorických neuronů. Dualita fusimotorického účinku byla pozorována již od
počátku, neboť se zdálo, že účinek jednotlivého gama-motoneuronu je buď čistě
statický nebo dynamický. Za náhodnou nepovažujeme spojení mezi jednotlivými
fusimotorickými neurony a jedním ze dvou typů fusimotorických účinků. Proto
hovoříme o dvou důležitých důsledcích (Hulliger, 1983; Matthews, 2000):
1.
Během
vývoje může docházet k vedení jednotlivých rozbíhajících se větví
fusimotorických neuronů k jejich specifickým intrafusálním cílům. Je také
možné, že fusimotorické neurony, poté co vytvoří funkční kontakty
s intrafusálními vlákny, by mohly determinovat diferenciaci těchto vláken
buď na bag nebo chain vlákna.
2.
Nositeli
specifických statických či dynamických vlastností jsou jednotlivé fusimotorické
neurony. Statické a dynamické vlastnosti by mohly být přístupné selektivní
nervové kontrole ze strany periferních reflexů nebo sestupných drah
z vyšších motorických center.
Nicméně od počátku byla rozlišovacím znakem dynamického účinku
skutečnost, že během svalového prodloužení se jedná o silný účinek na rozdíl od
jeho malých účinků během zkrácení svalu, umožňující klid aferentních vláken
během pomalého uvolnění. Na druhou stranu statická fusimotorická vlákna
normálně udržují aferentní výboj během svalového zkrácení (Hulliger, 1983).
Sekundární aferentní vlákna normálně působí jen přes statické
fusimotorické motoneurony. V typickém případě vytvářejí statická vlákna
prodloužené dráždivé účinky a mohou zvyšovat statickou polohovou citlivost,
která je doprovázená menšími účinky na dynamickou citlivost než u primárních
aferentních vláken (Hulliger, 1983).
2. 7 Fusimotorické účinky
Odpovědi primárních i
sekundárních aferentních zakončení na změnu protažení receptoru nazýváme „statická“ odpověď. Pokud je
receptorická část svalového vřeténka natažena pomalu, pak počet impulsů přenášených
jak z primárních, tak i sekundárních zakončení stoupá téměř přímo
úměrně se stupněm natažení a po
zakončení pokračují v přenášení těchto
impulsů po několik minut. Tento účinek
je nazýván statickou odpovědí vřeténkového receptoru, což vyjadřuje prostě to,
že jestliže zůstává samotný receptor
natažen, pak jak primární, tak i sekundární zakončení pokračují
v přenosu svých signálů po několik minut. Protože nuclear chain intrafusální vlákna jsou inervována jak
primárními, tak i sekundárními
zakončeními, má se za to, že tato nuclear chain vlákna jsou
nejvíce zodpovědná za statickou
odpověď (Boyd, 1998, Guyton, 2000; Hulliger, 1983).
Odpověď primárních (ale ne sekundárních) zakončení na míru změny délky receptoru nazýváme „dynamická“
odpověď. Pokud
se délka vřeténkového receptoru zvýší náhle, pak jsou primární zakončení
(ale ne sekundární zakončení) stimulována
obzvláště silně, mnohem silněji než při stimulaci zapříčiněné statickou odpovědí.
Tento zvýšený stimulus primárních zakončení je nazýván dynamickou odpovědí, což znamená, že
primární zakončení odpovídají extrémně
aktivně na rychlou míru změny délky vřeténka. Pokud dojde ke zvýšení
délky ve zlomcích sekund pak, dokonce i když
se délka vřeténkového receptoru zvýší
jen o zlomek mikrometru, přenášejí primární receptory ohromné
množství impulsů do Ia vláken, což se
ale děje jen v době, kdy dochází ke skutečnému zvětšování délky. Ihned po
zastavení zvětšování délky se míra impulsních výbojů rychle vrací na úroveň
mnohem menší statické odpovědi, která je stále v signálu přítomna
(Atwood, 1989; Enoka, 1994; Trojan a kol., 1996).
Naopak, když dochází ke zkracování vřeténkového receptoru, zapříčiňuje tato změna snížení výkonu impulsů z primárních zakončení; když dosáhne receptorová oblast své nové zkrácené
délky, dochází k znovuobjevení
impulsů v Ia vláknech, a to ve zlomcích sekund. Takže primární
zakončení posílají extrémně silné, buď positivní nebo negativní, signály
do spinální míchy, aby ji seznámily s jakoukoliv změnou délky v
receptorové oblasti vřeténka. Protože dynamickou odpověď přenášejí jen primární zakončení a téměř všechna nuclear bag intrafusální
vlákna mají jen primární zakončení,
předpokládá se, že nuclear bag
vlákna jsou zodpovědná za silnou dynamickou odpověď (Hulliger,
1983; Trojan a kol., 1996).
Normálně, zvláště pokud je přítomno menší množství podráždění
gama neuronů, vysílá svalové vřeténko impulsy senzomotorickým nervům
nepřetržitě. Svalová vřeténka mohou tedy posílat do spinální míchy buď
pozitivní signály – tzn. zvýšený počet impulsů naznačující zvyšující se
natažení svalu nebo negativní signály – počet impulsů snížený pod normální
úroveň, naznačující, že sval je uvolňován. Mezi účinky vyvolávanými
fusimotorickými neurony existuje vysoká míra variability. Jak velikost
dráždivého účinku (změna ve statickém pálení nebo fusimotorická chyba), tak i
množství změn v dynamické citlivosti (natažení svalového vřeténka zvyšuje
míru pálení výbojů, zatímco zkrácení míru pálení snižuje), může dosahovat širokého rozpětí. Později byl
přehodnocen stupeň čistoty duality a byl rozdělen do šesti skupin: od čistě
dynamických (I) až po čistě statické (VI). Původní duální rozdělení je stále
poměrně blízké pravdě protože 2/3 účinků byly buď čistě dynamické nebo čistě
statické. Většina statických axonů je selektivní a důsledně inervuje buď jen
„bag“2 nebo „chain“ vlákna. Vzhledem k tomuto faktu by „bag“2 vlákna mohla
být selektivně kontrolována z mozkové kůry (Bakker, 1982; Trojan a kol., 1996).
2. 7. 1 Kontrola intenzity statické a dynamické odpovědi gama
motorickými nervy
Gama motorické nervy
vedoucí k svalovému vřeténku mohou být rozdělena na dva typy:
gama-dynamické (gama-d) a
gama-statické (gama-s) nervy. První z nich dráždí hlavně „nuclear bag“ intrafusální vlákna a druhý
typ hlavně nuclear chain vlákna. Když
gama-d vlákna podráždí „nuclear bag“
vlákna, dochází k ohromnému zesílení
dynamické odpovědi svalového vřeténka, zatímco statická odpověď je jen málo ovlivněna. Naopak stimulace gama-s vláken, jež
dráždí „nuclear chain“ vlákna zesiluje
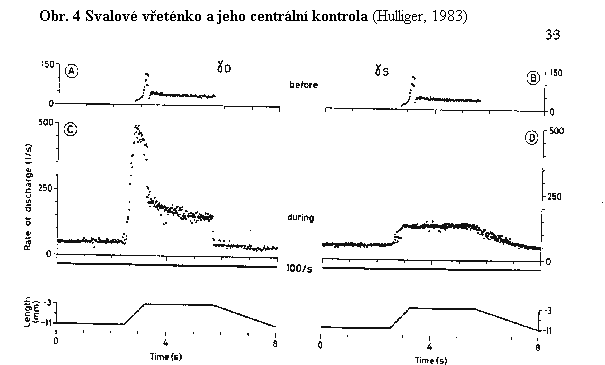
statickou odpověď zatímco dynamická odpověď je tímto jen velmi málo ovlivněna, viz. str. 26, obrázek 4
(Hulliger, 1983).
![]()
Vysvětlivky k obrázku 4:
before – před
during –
v průběhu
length –
délka
rate of
discharge (1/s) – měřítko výkonu
Popis obrázku
4: A - D – chování
dynamických a statických
fusimotorických
neuronů při
podráždění primárního aferentu za
stahu a
udržení
izometrické kontrakce
A a B – pasivní kontrola fusimotorické stimulace
C
- odpověď v průběhu tetanické
stimulace dynamického
gama
axonu při 100/s
D – odpověď v průběhu tetanické
stimulace statického gama
axonu
při 100/s
2. 8 Síla a rychlost fusimotorické akce
Síla a rychlost patří mezi
důležité parametry pro jakékoli hodnocení fusimotorických neuronů
v motorické kontrole.
Síla – v dřívějších studiích byly statické a dynamické gama-motoneurony
stejně účinné při ovlivňování primárních aferentních vláken na změnách kratších
svalových délek. Při současných měřeních na změnách delších svalových délek
však byly statické axony více než dvakrát silnější při dráždění Ia primárních
aferentních vláken než dynamické axony. Rozdíl mezi těmito údaji odráží odlišný
účinek na polohovou citlivost, která má tendenci se zvyšovat účinkem statických
a snižovat účinkem dynamických vláken. Funkčně se zdá být tento rozdíl
v síle dráždění významnější, protože jednotlivá vřeténka jsou normálně
inervována 2 – 3 krát větším množstvím statických než dynamických fusimotorických
vláken. Dynamické fusimotorické neurony se díky schopnosti zvýrazňovat
citlivost, zdají být vhodné pro kontrolu dynamické citlivosti. Oproti tomu
statický fusimotorický systém se zdá být zvláště účinným nástrojem pro kontrolu
celkové úrovně výboje vřeténkových aferentních vláken. Někdy i jediný impuls
působící na jediné gama axony nebo beta vlákna během dynamického natažení,
vyvolá silné podráždění Ia aferentních vláken, způsobené pravděpodobně zvýšenou
polární tuhostí bag1 vláken (Guyton, 2000; Hulliger, 1983).
Rychlost – dynamická fusimotorická akce byla dlouho považována
za značně pomalejší než statická. Tento názor vysvětluje nápadný rozdíl účinků
obou akcí na dynamickou citlivost primárních aferentních vláken, které lze
pozorovat u obojživelníků. Díky jejich vysoké míře změn mostů (mezi vlákny
aktinu a myosinu) by rychle se stahující vlákna měla být schopna pohlcovat
hlavní podíl dynamických délkových změn, bez toho, aby vytvářela tzv. dynamické
napěťové přechody. Mosty by měly být připoutány jen na tak krátkou dobu, která
znemožňuje jejich účinné natažení. U savčího vřeténka se zdá být rozdíl
rychlosti stahu mezi bag1 a bag2 nebo chain vlákny příliš malý na to, aby
vysvětlil velké rozdíly mezi dynamickým a statickým fusimotorickým účinkem na Ia
citlivost. Na úrovni intrafusálních vláken, byly nalezeny relativně malé
rozdíly v rychlosti stahu. U statické akce je dvakrát vyšší než u akce
dynamické (Guyton, 2000; Hulliger, 1983; Trojan a kol., 1996).
2. 9 Myotatický reflex
Nejjednodušší ukázkou funkce
svalového vřeténka je reflex svalového
natažení (také nazývaný myotatický reflex) – což znamená, že kdykoliv je sval náhle natažen, podráždění vřetének způsobí reflexní stažení velkých kosterních svalových vláken stejného
svalu a také blízce spojených synergistických svalů (Atwood, 1989; Holubová,
2002; Trojan a kol., 2001).
2. 9. 1 Nervový okruh natahovacího reflexu
Obr. 5 ukazuje základní okruh natahovacího reflexu svalového vřeténka a ukazuje proprioceptorické nervové vlákno typu Ia vycházející ze svalového vřeténka a vstupující do dorzálního kořenu spinální míchy. Na rozdíl od většiny ostatních nervových vláken vstupujících do míchy, vstupuje jedna větev tohoto nervu přímo do předního rohu šedé hmoty míšní a je synapticky spojena s motorickými nervy vystupujících z předních rohů míšních, jež posílají nervová vlákna zpět do stejného svalu, odkud pochází aferentní vlákno svalového vřeténka. Tedy je to monosynaptická dráha jež umožňuje reflexnímu signálu po podráždění vřeténka vrátit s v nejkratším možném čase zpět do svalu. Některá vlákna typu II ze sekundárních vřeténkových zakončení také končí monosynapticky na anteriorních motorických neuronech. Nicméně většina vláken typu II (stejně jako mnoho kolaterál z Ia vláken primárních zakončení) končí na mnohonásobných interneuronech v šedé spinální hmotě a ty na oplátku přenášejí k anteriorním motorickým neuronům více zpožděné signály nebo slouží jiným funkcím (Deursen, 1996; Enoka, 1994; Opavský, 2002; Trojan a kol., 1996).
Obr. 5 Řízení napětí svalu
pomocí svalového vřeténka a
Golgiho
šlachového tělíska
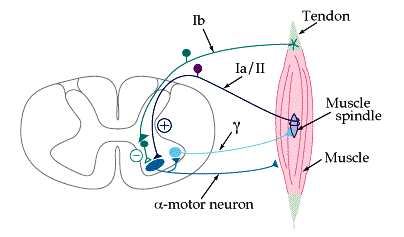
Vysvětlivky k obrázku 5: Tendon – šlacha
Muscle
spindle – svalové vřeténko
Muscle – sval
α - motoneuron – alfa motoneuron
2. 9. 2 Dynamický natahovací reflex versus statický natahovací reflex
Natahovací reflex může být rozdělen na dvě složky: dynamický natahovací reflex a statický natahovací reflex. Dynamický natahovací reflex je vyvolán silným dynamickým signálem přenášeným z primárních senzorických zakončení svalových vřetének, zapříčiněný rychlým natažením svalu. To znamená, že když je sval náhle natažen, je přenášen silný signál do spinální míchy a to způsobuje okamžitý silný reflexní stah stejného svalu, ze kterého signál pochází. Reflex tedy funguje jako nástroj čelící náhlým změnám délky svalu, protože svalový stah brání natažení. Dynamický natahovací reflex zaniká ve zlomcích sekund poté co je sval natažen do své nové délky a po následující prodloužené období pokračuje jen slabší statický natahovací reflex. Tento reflex je vyvoláván nepřetržitými statickými receptorickými signály přenášenými jak primárními, tak i sekundárními zakončeními. Důležitost statického natahovacího reflexu tkví v tom, že pokračuje ve vyvolávání svalového stahu po alespoň několik sekund nebo minut, kdy je sval udržován v nepřiměřené délce (Boyd, 1998; Hulliger, 1983; Hunt, 1990).
2. 9. 3 Negativní natahovací
reflex
Když je sval náhle zkrácen, dochází k přesně opačnému
efektu díky sníženému počtu impulsů
z vřetének. Pokud je sval již napnut, pak náhlé uvolnění
zatížení, jež je na sval kladeno, umožní zkrácení svalu a vyvolá jak dynamický, tak i statický
reflex svalové inhibice než svalové podráždění. Negativní natahovací reflex
tedy brání zkrácení svalu stejným způsobem jako positivní natahovací
reflex brání prodloužení svalu. Můžeme tedy vidět, že natahovací reflex má tendenci udržovat stálou délku svalu
(Guyton & Hall, 2000; Hulliger, 1983; Matthews, 2000).
2. 9. 4 Tlumící funkce dynamických a statických natahovacích reflexů
Zvláště důležitou funkcí
natahovacího reflexu je jeho
schopnost bránit oscilaci nebo trhavosti tělesných pohybů. To je tzv. tlumící neboli
zjemňující funkce. Např. použití
tlumícího mechanismu při zjemnění svalového stahu - signály
z oblastí spinální míchy a jiných částí
nervového systému jsou často přenášeny ke svalu v nezjemnělé formě,
na několik milisekund zvyšuje svou
intensitu, pak ji zase snižuje, pak ji znovu mění na jinou úroveň a tak dále.
Když vřeténkový aparát svalu nefunguje
dostatečně, je svalový stah během takovýchto signálů trhavý (Hulliger, 1983;
Chaitow, 1996; Trojan a kol., 1996; Trojan a kol., 2001).
2. 10 Role svalového vřeténka při volní motorické aktivitě
Abychom zdůraznili důležitost
gama eferentního systému musíme si uvědomit že 31% všech motorických
nervových vláken vedoucích ke svalu je
gama eferetních vláken a ne typu alfa motorických vláken. Kdykoliv je signál přenášen z motorické
kůry nebo z jakékoliv jiné
oblasti mozku k alfa motorickým
neuronům, jsou ve většině případů ve stejné době stimulovány také gama
motorické neurony, což je jev nazývaný ko-aktivace alfa a gama motorických
neuronů. To zapříčiňuje, že extrafusální a intrafusální svalová vlákna se
stahují ve stejné době. Smyslem stáhnou svalové vřeténko ve stejné době jako
větší kosterní svalová vlákna je: za prvé udržení délky receptorické části
svalového vřeténka beze změny, a tak
znemožnit svalovému vřeténku bránit
svalovému stahu. A za druhé udržuje vlastní tlumící funkci svalového
vřeténka bez ohledu na změnu délky
svalu. Například, jestliže by nedošlo ke stažení a uvolnění svalového vřeténka
spolu se svalovými vlákny, pak by receptorická
část vřeténka mohla být někdy
příliš uvolněná a jindy příliš natažená, což by v obou případech vedlo
jen k tomu, že by fungovalo
v jiných než optimálních podmínkách (Atwood, 1989; Enoka, 1994; Guyton
& Hall, 2000; Hulliger, 1983).
2. 10. 1 Stabilizace svalového vřeténka v průběhu napínací akce
Jedna z nejdůležitějších funkcí systému svalového vřeténka
je stabilizovat pozici těla během napínací motorické akce. Aby tuto
funkci splnily, přenášejí bulboretikulární facilitatorní oblasti a s ní
spojené oblasti mozkového kmene přes gama nervová vlákna dráždivé signály do intrafusálních svalových
vláken svalových vřetének. To zkracuje
koncové části vřetének a natahuje
centrální receptorické oblasti, a tak zvyšuje jejich signální výstup.
Nicméně vřeténka na obou stranách
každého kloubu jsou aktivována ve stejnou dobu. Proto
se také reflexní podráždění kosterních svalů zvyšuje na obou stranách
kloubu a vytvářejí se tak utažené
a napjaté svaly navzájem si v kloubu protiléhající. Čistým výsledkem je, že se pozice kloubu
silně stabilizuje a jakákoliv síla, která má tendenci pohnout s kloubem z jeho současné pozice, čelí vysoce citlivým natahovacím reflexům působícím na obou stranách kloubu. Kdykoli
musí osoba provést svalovou funkci, jež požaduje vysoký stupeň jemné a přesné pozice, stabilizuje
podráždění příslušných svalových
vřetének signály
z bulboretikulární facilitatorní oblasti mozkového kmene pozice hlavních
kloubů. To ohromně pomáhá
v provádění doplňkových
volních pohybů (prstů nebo jiných tělních částí) potřebných pro složité
motorické procedury (Boyd, 1998; Holubová, 2002; Hunt, 1990; Opavský, 2002).
2. 11 Mozkové oblasti kontroly gama-motorického systému
Gama eferentní systém je drážděn zvláště signály z bulboretikulární facilitatorní
oblasti mozkového kmene a sekundárně
impulsy přenášenými do
bulboretikulární oblasti z 1) mozečku, 2) basálních ganglií a dokonce i z 3) mozkové kůry.
Bulboretikulární facilitatorní oblast je zvláště spojena s antigravitačními
svaly a protože antigravitační svaly
mají zvláště velkou hustotu
svalových vřetének, je zdůrazňována
důležitost gama eferentního mechanismu
v tlumících pohybech různých
částí těla během chůze a běhu
(Atwood, 1989; Holubová, 2002; Guyton & Hall, 2000; Trojan a kol., 1996;
Véle, 1996).
2. 12 Klinické aplikace myotatické reflexů
Cílem tohoto jednání je určit
jak mnoho podráždění na pozadí
neboli „tonu“ vysílá mozek do spinální míchy. Tento reflex je vyvoláván následujícím způsobem (Atwood,
1989).
2. 12. 1 Patelární reflex a jiné reflexy
Patelární reflex může být vyvolán jednoduše úderem neurologického kladívka na patelární šlachu, to
způsobí protažení čtyřhlavého svalu a
podráždí svalová vřeténka, která pak způsobí, že se spustí dynamický natahovací
reflex, noha vykopne dopředu, viz str. 33, obrázek 6. Podobné reflexy mohou být vyvolány téměř u všech svalů
těla a to buď úderem na šlachu
svalu nebo úderem na svalové bříško.
Jinými slovy náhlé natažení svalových vřetének, je vše co je nutné pro vyvolání natahovacího reflexu. Svalové reflexy jsou
neurology používány k odhadnutí
výše postižení center spinální míchy. Když je do míchy z horních oblastí centrálního nervového systému přenášen velký počet impulsů
jsou svalové reflexy velmi bolestivé a reflexní zóna je rozšířena
(hyperreflexie). Naopak, jestliže jsou
impulsy potlačeny nebo odstraněny, jsou svalové reflexy značně oslabené nebo úplně chybí (hyporeflexie,
popř. areflexie). Tyto reflexy jsou často používány při výskytu poškození
v motorických oblastech mozku nebo
při určování nemocí, jež dráždí
bulboretikulární facilitatorní oblast mozkového kmene. Obyčejně velká poškození v motorických oblastech mozkové kůry, zvláště ty, které byly způsobeny nárazem nebo mozkovým nádorem, jsou velmi
bolestivé (Holubová, 2002; Opavský, 2002; Trojan a kol., 2001).
Obr. 6 Záznam
EMG z musculus quadriceps femoris v průběhu vyvolání
patelárního reflexu (nahoře) a musculus gastrocnemius v průběhu
vyvolání achilárního reflexu
(dole)
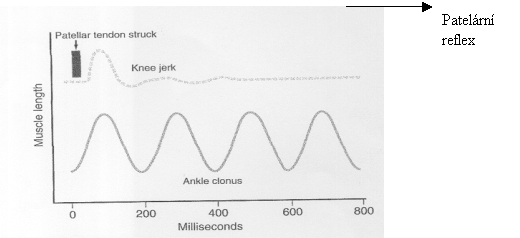
Vysvětlivky k obrázku 6: Ankle clonus – klonus kotníku
Patellar tendon struck – úder na
patelární šlachu
Knee jerk – patelární reflex
Muscle length – svalová délka
Milliseconds - milisekundy
2. 12. 2 Klonus
Při vhodných podmínkách může svalový reflex oscilovat – jev
zvaný klonus, viz. str. 34, obrázek 7. Oscilace může být vysvětlena zvláště dobře ve vztahu k bicipitovým
klonusům – viz níže (Hulliger, 1983).
Obr. 7 Svalová kontrakce způsobená spinálním signálem za dvou podmínek:
křivka A v normálním svalu a křivka B ve svalu denervovaném
(Hulliger,
1983)
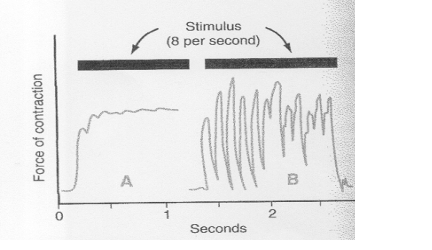
Vysvětlivky
k obrázku 7: Stimulu (8 per second) – stimul (po dobu 8 s.)
Seconds – sekundy
Force
of contraction – síla úderu
Jestliže osoba stojící na špičkách prstů náhle spustí své tělo
směrem dolů a natáhne tak jeden ze svalů musculus gastrocnemius, jsou přenášeny
impulsy z svalových vřetének do spinální míchy. Impulsy reflexně
podráždí natažený sval, který znovu vyzvedne tělo nahoru. Po zlomku
vteřiny reflexní stah svalu vymizí a tělo znovu spadne dolů a
tak natáhne vřeténka podruhé.
Dynamický natahovací reflex znovu
vyzvedne tělo, ale také ve zlomcích
vteřiny vymizí a tělo znovu spadne dolů
a vyvolá tak další cyklus. Tímto způsobem natahovací reflex svalu musculus
gastrocnemius pokračuje v oscilaci, často po dlouhé období – jev zvaný
klonus. Klonus se běžně vyskytuje je-li
natahovací reflex vysoce zcitlivěn facilitujícími impulsy z mozku. Např. u
decerebrovaného zvířete, u kterého jsou
natahovací reflexy velmi
usnadněny, se klonus vyvíjí rychle (viz patologie na obrázku č. 7, str. 34).
Neurologové testují pacienty na klonus náhlým natažením svalu, za udržování stálé natahovací síly uplatňované na sval proto, aby určili stupeň poškození spinální míchy. Jestliže
dojde ke záškubu, pak je jisté, že
stupeň poškození je vysoký (Boyd, 1998; Hulliger, 1983).
3 REHABILITACE
3. 1 Neurofyziologický podklad svalového tonu a hypertonu
Řízení svalové činnosti je uskutečňováno nervovou soustavou
prostřednictvím nervovým drah. Dráha gama-systému nastavuje práh dráždivosti
motoneuronů, zatím co dráha alfa-systému spouští aktivitu motoneuronu při
pohybu. Ve spánku je tonus nižší než v bdělém stavu, neboť v noci
klesá aktivita gama-systému řízeného retikulární formací. Gama-systém tedy
můžeme považovat za jeden ze systémů, který se podílí na regulaci svalového
tonusu (Holubová, 2002).
3. 2 Dělení svalového hypertonu
Svalový hypertonu je možno
dle etiologie rozdělit:
1. organická příčina – a. spasticita
Spasticitu lze vzhledem k její různorodosti jen obtížně definovat. Nejvíce je dnes přijímána definice Lanceho, dle níž je spasticita: motorická porucha, projevující se zesílením tonických napínacích reflexů, proprioceptivních šlachových reflexů, v závislosti na rychlosti protažení svalu; je podmíněna zvýšenou excitabilitou těchto reflexů, která je součástí centrální parézy (Pavlů, 1996).
Jak již bylo řečeno, jedním ze základních rysů spasticity je zvýšení svalového tonu závislé na rychlosti spolu s vystupňovanými myotatickými reflexy. I když základní reflexní oblouk z primárních zakončení vřeténka vlákny Ia k synergickému svalu je monosynaptický, regulace napínacího reflexu se uskutečňuje oligosynaptickými a polysynaptickými okruhy (Mayer, 1997).
Spasticita je častým průvodním příznakem při lézích prvního motoneuronu. Příčinou nejspíše není jeho přímé poškození, ale jsou zasaženy extrapyramidové dráhy, které probíhají v jeho bezprostřední blízkosti a mají inhibiční vliv na periferní motoneuron. Na světové konferenci o dětské mozkové obrně (DMO) v Edinburgu v roce 1969 byla spasticita definována jako „trvalé zvýšení napínacích reflexů, vedoucí k nárůstu svalového odporu k napínání. Napětí může náhle poklesnout, známý fenomén sklapovacího nože“ (Mayer & Konečný, 1998; Seidel, Obenberg & Süssová, 2002).
2. funkční
příčina
(Seidel, Obenberg & Süssová, 2002)):
a.
dysfunkce limbického systému
– svaly nejsou spontánně bolestivé, ale jsou
citlivé na pohmat. Projevem nedostatečné relaxace svalu je spontánní klidová
aktivita na elektromyografii (EMG). V závislosti na poloze se mění hranice
mezi hypertonickou a normotonickou oblastí, z čehož je patrný vliv
gama-systému. Ve vertikální poloze je rozsah hypertonu větší než vleže.
Nepostihuje izolované svaly, ale celé skupiny svalů.
b.
dysfunkce na úrovni míšního
segmentu – sval
je spontánně bolestivý a enormně citlivý na protažení, včetně pohmatu. Hlavním
činitelem jsou vmezeřené interneurony, charakteristické je postižení anatomicky
definovaného svalu. Není přítomna žádná klidová EMG aktivita. Antagonistický
sval (či skupina) je v inhibici, na základě reciproční inhibice.
c.
inkoordinace svalových
vláken – Na
podkladě funkční poruchy vzniká izolovaná porucha relaxace (dekontrakce)
několika svalových vláken označovaná jako vnitřní inkoordinace. Kontrakce
svalových vláken způsobuje permanentní tah za úpon svalu a je nejčastější
příčinou entezopatíí. Trvale kontrahovaná vlákna jsou ischemizována, tím pádem
mohou vznikající metabolity dráždit nociceptory.
3. 3 Mechanismus vzniku spasticity
1. Teorie zvýšené aktivace gama-motoneuronů – centrálně vyvolané
zvýšení aktivity gama-motoneuronů zvyšuje citlivost svalových vřetének, což
vede ke zvýšení napínacích reflexů.
2. Imbalanční teorie – u korových lézí dochází k převaze
tonicko-exitačních sestupných drah, což vede ke zvýšení dráždivosti míšních
alfa-motoneuronů, a tím ke zvýšení svalového tonu
3. Teorie „sproutingu“ – Výpadem sestupných axonů, způsobených
centrálních lézí, se uvolní na alfa-motoneuronu synaptická místa, která se pak
obsazují segmentálními exitačními aferencemi. Ke zvýšení tonu tedy dochází
posunem rovnováhy směrem k exitaci. Tato teorie byla však pomocí
elektronové mikroskopie silně zpochybněna.
3. 4 Mechanismus zapojení svalového vřeténka v průběhu pasivního
pohybu
Pokud protahujeme sval
pasivně, postupně se v určitém okamžiku začíná vyvíjet odpor proti tomuto
pohybu. A tak brání volnému pohybu v kloubu. Tato překážka je vyvolána
pozvolným vzrůstem napětí v měkkých tkáních kolem kloubu při zvětšování
rozsahu pohybu. Největší vliv na vznik tohoto napětí má svalová tkáň. Velikost
tohoto vyvolaného napětí závisí na úhlu rotace v kloubu, který ovlivňuje
délku svalu a tím i jeho excitabilitu. Prodlužování svalu zvyšuje excitabilitu
svalu a tím vyvolává tendenci k jeho zkracování na výchozí nastavenou
délku. Intenzita této tendence k odporu proti natahování je závislá i na
rychlosti, s jakou se pohyb provádí. Při pomalé rychlosti pohybu
v kloubu lze tuto svalovou reakci vnímat jako pružný růst napětí svalu při
jeho protahování. To může vznikat jednak velmi sporadickou asynchronní aktivací
některých dráždivějších motorických jednotek nebo i aktivací intrafusálních
kontraktilních vláken při protrakci svalového vřeténka. Toto napětí vzrůstá i
se zvyšující se rychlostí protažení, až při značné rychlosti přejde toto
zvyšování napětí do synchronního reflexního klonu, který označujeme jako
elementární posturální reflex (ERP) (Božíková, 1999; Véle, 1996).
Pasivní protahování svalů se zdá být velmi důležitou složkou
tlumení spasticity. Jestliže sval dlouho není protahován, dochází ke snížení
viskoelasticity svalových vláken, což zvyšuje excitabilitu svalových vřetének
(Pavlů, 1999).
3. 5 Koncepty, metody a přístupy používané ve fyzioterapii
k ovlivňování hypertonu a
spasticity
3. 5. 1 Koncept manželů Bobathových
Je indikován pro motorické poruchy v důsledku DMO, ale také
v důsledku hemiplégie dospělých. Podstatou tohoto konceptu je tlumení
spasticity.
Hlavními terapeutickými prostředky jsou uvádění těla či
jednotlivých segmentů těla do tzv. antispastických poloh a setrvalé protahování
spastických svalů. To vede k adaptaci svalových vřetének na nové podmínky,
takže po návratu do výchozí polohy je aktivační účinnost aferentních impulsů
snížena. Důsledkem je také snížení aktivační pohotovosti motoneuronů a ubývání
spasticity. Koncept byl během několika desetiletí dále rozvíjen a stále více
zaměřován na běžné aktivity denního života (Pavlů, 1996; Pfeiffer a kol.,
1976).
3. 5. 2 PNF (proprioceptivní neuromuskulární facilitace)
Základním principem je cílené ovlivňování aktivity motorických
neuronů předních rohů míšních prostřednictvím aferentních impulsů ze svalových,
šlachových a kloubních proprioreceptorů.
PNF techniky pracují s postfacilitační inhibicí, kdy se
předpokládá, že po facilitaci svalu dochází k jeho útlumu.
Aplikace vytahovacích technik v útlumové fázi se dá
rozdělit do dvou základních typů:
·
vytažení svalu – týká se pouze jediného
svalu, se kterým právě pracujeme a pacient neaktivuje jinou svalovou skupinu
·
vytažení svalu s volní
aktivací antagonistické svalové skupiny - jednou z velkých předností antagonistů
je, že dochází k prohloubení k útlumu vytahovaného svalu. Nevýhodou
je, že volní kontrakce vede ke zvýšení dráždivosti celého svalového systému,
což může do určité míre postfacilitační útlum vyrušit. Přídavné volní aktivace
antagonisty proti vytahovanému svalu je indikována až tehdy, jeli pacient
schopen s námi plně spolupracovat.
Poslední
výzkumy ukazují, že tyto techniky prokazatelně působí pouze na svalové vlákno.
Intersticiální vazivo, šlachy a vazy pravděpodobně unikají možnosti
terapeutického ovlivnění. Dalším důležitým faktorem aplikace PNF technik je
optimální načasování a počet opakování vytahovacího manévru. Experimentální
výsledky ukazují, že plného postfacilitačního útlumu dosáhneme po maximální
svalové kontrakci trvající 1 s. Z toho vyplývá, že maximální svalová
kontrakce delší než 1 s nemá vliv na velikost svalové relaxace.
V praxi se ovšem setkáváme s kontrakcí trvající 5 – 10 s (dle toho,
zda se jedná o sval fázický či tonický). Sval je v útlumu ihned po
kontrakci po dobu 5 s. Protahovat musíme ihned po kontrakci. Jinak by mohlo
hrozit, že budeme protahovat sval, který již není v útlumu (Adler a kol.,
1993; Capko, 1998; Holubová, 2002; Pavlů, 1996; Pfeiffer a kol., 1976).
3. 5. 3 Postizometrická relaxace (PIR)
Metodu PIR řadíme do
kategorie neuromuskulární technik, které využívají v terapii
postfacilitačního útlumu. Je to prakticky nejpoužívanější technika při nálezech
zvýšeného svalového napětí se známkami reflexních změn. Charakterizuje ji
použití velmi malé síly při volní kontrakci tak, aby byla aktivována pouze
vlákna s TP (trigger points) (Vodičková, 1996).
Při terapii vycházíme ze svalového předpětí, ve kterém pacient
provádí izometrickou kontrakci svalu. Po fázi izometrické kontrakce čekáme na
spontánní uvolnění svalu. Úkolem je pouze vést pohyb v požadovaném směru,
neúčastníme se aktivní silou (neprotahujeme!). Síla, kterou během izometrické
kontrakce vyvíjí je minimální. Během odporu při minimální síle se aktivuje jen
malý počet vláken, zatímco většina zůstává utlumena. Tím jsou zasažena jen
vlákna spojená s TP, která jsou charakteristická nižším prahem dráždivost
vzhledem k okolním svalovým vláknům (Lewit, 1996).
Ve fázi izometrické kontrakce dosáhneme určitého metabolického
„chaosu“ na úrovni mikrostruktury svalu, a zároveň ovlivníme vyšší struktury
CNS. Ve fázi relaxace pak má sval dostatek času na úpravu vnitřních podmínek,
které byly aktivací rozbourány (Vodičková, 1996).
U reflexních změn ve svalech a obecně v hypertonickém
svalu je abnormálně zvýšené nastavení aktivity jak alfa, tak i
gama-motoneuronů. Toto vyšší nastavení gama-systému zvyšuje také aferentní
množství informací jdoucích ze svalových vřetének (Ia i II), což celý kruh
uzavírá a udržuje. Další okolností, která dovoluje tomuto okruhu fungovat, je
absence nebo snížení kortikální inhibice (Mitchell, 1993).
Jedou z možností, jak do tohoto kruhu vstoupit, je použití
izometrické kontrakce. Izometrická kontrakce nám pomůže změnit nastavení
gama-smyčky a ovlivnit míšní interneurony a suprasegmentální inhibiční synapse
(Mitchell, 1993).
Izometrická kontrakce extrafusálních vláken podle Michella
(1993) vyvolá protažení „nuclear bag fibers“, což je způsobeno posuny svalových
fibril při izometrické kontrakci vůči sobě navzájem (neprojeví se změnou délky
svalu). Toto „zevní“ protažení vyvolá krátkodobou excitaci aferentních vláken
svalového vřeténka, které se však ihned adaptují na nově dosaženou délku
intrafusálních vláken. Při izometrické svalové kontrakci tlačí extrafusální
vlákna na pouzdro svalového vřeténka, čímž ho zužují a zmenšují tak
subkapsulární lymfatický prostor, pročež dochází k vytlačování tekutiny
z tohoto prostoru, navíc se objevuje mírná redukce aferentních informací
jdoucích ze sekundárních aferentních zakončení (II). U primárních aferentních
zakončení dochází k rychlé adaptaci.
Ve fázi postizometrické relaxace mohou být extrafusální vlákna
protažena bez současného protažení „nuclear bag fibers“ a bez stimulace aferentních
informací ze svalových vřetének, což je umožněno novým nastavením
gama-eferentního systému (snížení frekvence impulsů vycházejících
z gama-motoneuronů) (Mitchell, 1993; Božíková, 1999).
Předpokládá se, že PIR může přeprogramovat i suprasegmentální
systémy. Volní izometrická kontrakce vyžaduje spolupráci pacienta a tudíž vede
k zapojení mozkové kůry, extrapyramidového a limbického systému do
léčebného procesu (Mitchell, 1993).
Při této metodě je nutno vyčkat spontánního uvolnění, neboť
pokud se izometrickou kontrakcí nepodaří snížit aktivitu gama-eferentního
systému, pak nelze dosáhnout potřebné relaxace. Jestliže protahujeme sval
aktivně bez relaxace, dochází ke zvyšování aktivity gama-systému, tím dojde ke
zvýšenému množství aferentních informací ze svalových vřetýnek a tudíž ještě
k silnější reflexní svalové kontrakci (Smékal, 1998).
3. 5. 4 Metoda excentrické dekontrakce (ED) dle Brüggerova konceptu
Metodu
ED řadíme do kategorie neuromuskulárních technik, které využívají
v terapii recipročního útlumu. Někdy označujeme takové techniky jako
antagonistické.
ED využívá aktivní excentrické kontrakce svalu, který má
antagonistickou funkci vzhledem ke svalu, který má být terapií ovlivněn
(agonista).
Z neurofyziologického
pohledu je mohutnost proprioreceptivní aferentace z příslušných svalů
v průběhu excentrické dekontrakce vyšší než u koncentrické, protože jsou
nadprahově podrážděna jak svalová vřeténka (sval se prodlužuje), tak Golgiho
tělíska (sval vyvíjí tenzi, a proto stoupá napětí na rozhraní svalové a
šlachové části). Mohutnější aferentace vede ke snížení prahu dráždivosti
alfa-motoneuronů, a proto některé studie uvádějí excentrickou dekontrakci jako
kontrakci s nejmohutnějším náborem motorických jednotek. Kineziologicky
lze charakterizovat excentrickou dekontrakci jako brzdní pohyb ve směru
působení gravitace. V rámci nerušeného, harmonicky probíhajícího
„fyziologického“ pohybu spolupracují všechny svaly, respektive svalové smyčky
„synergisticky“. Při koncentrických kontrakcích agonisty se antagonista účastní
pohybu v kloubu prostřednictvím excentrických kontrakcí „dekontrakcí“, což
je princip recipročního útlumu (Brügger, 1986; Havlíčková, 1999).
Tato metoda zahrnuje úpravu
poruchy svalové dekontrakce u hypertonického svalu, který se projevuje svalovým
zkrácením. Je důležité, aby při návratu z konečné polohy nedošlo
k přerušení dosaženého snížení toku aferentních informací rychlým návratem
(s rychlým protažením intrafusálních vláken) svalu do počáteční polohy. Zkrácený
sval tedy učíme znovuzapojení do funkčního pohybu tak, aby výsledkem byl
koordinovaný pohyb (Kusunuse, 1993; Vodičková, 1996).
Možnou nevýhodou metody ED je vlastní provádění excentrické
dekontrakce, která musí být koordinovaná, což vyžaduje přesnou souhru (sladění)
mezi silou vyvíjenou terapeutem a silou, kterou pacient provádí kontrakci svalu
(Smékal, 1998).
3. 5. 5 Metoda dle sestry Kenny
Byla vypracována pro léčení
dětské obrny, poliomyelitis anterior acuta. Skládá se z horkých zábalů
zmírňujících bolest, uvolňujících tkáň a z pasivního vytahování
zkracujících se tkání, polohování a nácvik volních pohybů. Ten se koná
analyticky u každého svalu. Proprioreceptory se dráždí ruční vibrací, tření
šlach a dotekem kůže nad svalem. Stimulací je chvějivý pohyb, který facilituje
zakončení senzorických nervů v kloubu, šlachách a ve svalu. Sval je
v první fázi protažen, což vyvolá mohutných příliv dostředivých podnětů
z periferie, zejména ze svalových proprioreceptorů. Napětí extrafusálních
vláken agonisty se zvýší protažením a při sakadovaných pohybech, kdy dochází ke
zkracování délky svalu musí být zároveň přenastavováno napětí intrafusálních
vláken pomocí gama-moteneuronů z předních rohů míšních. Zároveň jsou tlumeny prostřednictvím
interneuronu alfa-motoneurony antagonistů. V interneuronech se facilituje
nebo tlumí základní aktivita, hovoříme o reciproční inervaci. Zároveň je třeba
se snažit, aby se nervové popudy nedostávaly k nesprávným svalů a
nevznikaly tak svalové inkoordinace. Cílem je přesná funkce a nikoliv síla (Pfeiffer a kol., 1976).
4 DISKUSE
Struktura svalového vřeténka je v podstatě známa a
jednotliví autoři se na ni v zásadě shodují.
Svalové vřeténko je specializovaný svalový receptor, který
informuje CNS jak o rychlých (fazických) změnách délky svalu (při pohybu), tak
i o změnách dlouhodobých, tonických (při udržování polohy)(Trojan, 1996).
Podle Kusunose (1993) je počet svalových vřetének ve svalech
fazických větší než ve svalech posturálních, což můžeme vysvětlit nutností
přesné a cílené kontroly při fazickém pohybu.
Velice málo je v literatuře hovořeno o tzv. statických a dynamických fusimotorických neuronech. Při dynamických pohybech (prudké natažení) svalového vlákna dojde k nastimulování Ia aferentních vláken a přes reflexní oblouk zpětně k nastavení svalového vřeténka pomocí gama-dynamických vláken. Gama-dynamická vlákna dráždí hlavně „nuclear bag“ intrafusální vlákna. Naopak statická fusimotorická vlákna normálně udržují aferentní výboj během svalového zkrácení a dráždí „nuclear chain“ vlákna (Hulliger, 1983).
Dnes již není žádnou novinkou, že během vývoje může docházet k vedení jednotlivých rozbíhajících se větví fusimotorických neuronů k jejich specifickým intrafusálním cílům. Také fusimotorické neurony, poté co vytvoří funkční kontakty s intrafusálními vlákny determinují diferenciaci těchto vláken buď na bag nebo chain vlákna.
Všechny výše zmíněné fyzioterapeutické koncepty a metody vykazují v početných případech viditelné až spektakulární úspěchy, týkající se zejména zlepšování centrálních poruch hybnosti u dětí i dospělých nebo pokroku motorického vývoje u dětí. Výsledky léčby však závisejí spíše na individuální vhodnosti metody vzhledem k charakteru postižení, ke spolupráci pacienta i k jeho rodinnému prostředí. Velký význam má při tom také motivace pacienta a jeho rodinných příslušníků. Dále pak velmi záleží na angažovanosti a speciálních schopnostech terapeuta, k průběžnému posuzování abnormálních držení a pohybů pacienta a jeho reakcí na jednotlivé fyzioterapeutické úkony, na schopnosti dostatečné míry empatie a pochopení klientova problému. Terapeut tedy musí zvládat nejen onu „praktickou“ stránku výkonu, ale musí být schopen vyřešit i řadu psychických problému pacienta. Z vlastní zkušenosti vím, že téměř žádné zdravotnické zařízení u nás nedokáže poskytnout nemocnému adekvátní psychologickou pomoc. Nemyslím si, že chyba je v již zmíněných zařízeních, dle mého názoru ma vinu na současném stavu systém zdravotnictví jako celku.
Jako negativní aspekty se však nezřídka objevuje dogmatický charakter některých konceptů a setrvávání na zastaralých či překonaných tvrzeních. Dalšími negativními jevy jsou proklamace univerzálních indikací a výlučnosti úspěchu, spojené se zavrhováním jiných metod. Původce některých metod a hlavně jejich následovníci vytvořili organizace, někdy i sektářského charakteru, které si osobují právo určovat jediný správný způsob provádění daných fyzioterapeutických úkonů a dále pak i výhradní právo ke školení odborníků. Za zcela zbytečné je dnes možno, v souladu s řadou kompetentních odborníků, považovat některé spory mezi zatvrzelými zástupci jednotlivých konceptů. Jako klasické příklady lze uvést kontroverze konceptů Vojta kontra Bobath, ale i Bobath kontra PNF. Z celé řady dosud provedených srovnání diskutovaných speciálních fyzioterapeutických metod lze usuzovat na to, že žádná z nich není co do dlouhodobého efektu v obecnějším smyslu jednoznačně nadřazena metodám ostatním.
Mezi jedny z nejvíce používaných metod v rehabilitaci
patří PIR, PNF a ED. Technika PIR je nejpoužívanější technika při nálezech
zvýšeného svalového napětí se známkami reflexních změn. Metoda PNF pracuje
s posfacilitační inhibicí, kdy předpokládáme, že po facilitaci svalu
dochází k jeho útlumu. Metodu ED řadíme mezi neuromuskulární techniky
využívající recipročního útlumu. Podle Vodičkové (1996) nebyl při metodách PIR
a ED použitých k ošetřování ischiokrurálního svalstva pacientů ve výsledném
efektu rozdíl a dokonce při kontrolních měřeních ischiokrurálního svalstva na
zdravých končetinách bylo patrné zlepšení. To svědčí o centrálním řízení
svalového systému.
Jednotlivé metody se od sebe vzájemně liší různým provedením,
tedy v podstatě se snažíme rozdílnými terapeutickými cestami dospět ke
stejným nebo podobným efektům léčby.
Metodou PIR indikujeme na svaly, u kterých najdeme jen několik
hypertonických svalových vláken. Charakteristické pro tuto metodu je použití
velmi malé síly při volní kontrakci tak, aby byla aktivována pouze vlákna
z TP. V průběhu odporu při minimální síle se aktivuje pouze malý
počet vláken, zatímco většina zůstává utlumena. Tímto mechanismem se zapojí
pouze vlákna s nižším prahem dráždivosti vzhledem k okolním svalovým
vláknům, tedy svalová vlákna, která jsou v hypertonu. Pokud aktivujeme jen
tato hypertonická vlákna dochází k „normální“ reakci svalových vřetének
v těchto vláknech. Dojde k protáhnutí intrafusalních vláken („nuclear
bag fibers“). Ty aktivují Ia a II nervová vlákna, která vedou informaci o změně
délky svalového vřeténka do míchy (přes zadní rohy míšní). Zde, dle mého
názoru, nastavají dvě možné odpovědi systému.
Za prvé dojde k aktivaci především gama-systému, který
přenastaví dané svalové vřeténko a umožní tak jeho opětovné podráždění.
Gama-systém tak při zachování nové délky svalového vlákna zajistí zkrácení
intrafusálních vláken svalového vřeténka a tím umožní jeho opětovnou
připravenost pro další podráždění svalového vřeténka. K tomu dochází
tehdy, pokud sumace vzruchů z Ia a II vláken nedosáhne úrovně potřebné
k podráždění alfa-motoneuronů. Tento způsob reakce se uplatňuje především
u postizometrické relaxace, kde nepatrnou svalovou kontrakcí, kterou můžeme
přirovnat k izometrické kontrakci, aktivujeme pouze hypertonická vlákna
daného svalu. Postupným opakováním této cílené aktivace hypertonických vláken
dosáhneme zrušení hypertonu.
Druhou možností odpovědí centra můžeme popsat principem
excentrické dekonztrakce (ED). Princip ED jsem vysvětlil v kapitole Rehabilitace. Neurofyziologický pohled
této metody je ovšem poněkud složitější. Myslím si, že pokud pacient zapojuje
agonisty, dochází k mohutné aferentaci pomocí Ia nervových vláken právě
těchto agonistů za současného zkracování hypertonických svalů fyzioterapeutem. Odpovědí
centra je masivní zapojení alfa-motoneuronu. Tím se snaží vrátit svalové vlákno
do výchozí polohy. Fyzioterapeut se však snaží zabránit svalu v provedení
„ochranné“ koncentrické kontrakce (fyzioterapeut „přetáhne“ pacienta). Současně
s tím dochází přepojením přes interneuron k inhibici antagonistů.
V okamžiku zastavení pohybu ustává bouřlivý přenos impulzu do Ia vláken a
následně i do centra. Pochopitelně ustává i zvýšená aktivita alfa-motoneuronu.
Následně dochází k aktivaci gama-systému, který přenastaví svalové
vřeténko na nově vzniklou situaci. Tím vlastně jsme dosáhli zmenšení hypertonu
svalu.
Vzhledem k tomu, že neurofyzilogické podklady těchto
metod nebyly zatím dokonale prostudovány, je nasnadě otázka, zda je primárním
cílem fyzioterapie zcela přesné pochopení těchto principů, nebo úspěch léčby a
spokojenost klienta.
5 ZÁVĚR
Cílem této bakalářské práce bylo ozřejmit a ucelit poznatky o svalovém vřeténko a jeho užití při některých rehabilitačních metodách.
1. První informace o svalovém vřeténku jako smyslovém orgánu byly známy z prací Sherringtona z roku 1894 a Ruffiniho z roku 1898. Až za poměrně dlouhou dobu byly připsány další informace o různých typech nervových zakončení vřeténka a jeho zapojení do centrálního řízení svalového tonu. V roce 1951 byly identifikovány gama-motoneurony pomocí rozdílných přenosových axonálních rychlostí.
2. Funkce uvědomování si polohy končetin vůči ostatnímu tělu i okolnímu prostředí je zajišťováno souborem fyziologických mechanismů. Mezi ně patří přenášení informací o svalovém tonu jednotlivých svalů z proprioreceptorů (svalových vřetének a Golgiho svalových tělísek). Svalová vřeténka jsou rozeseta na povrchu nebo uvnitř svalového bříška a posílají informace do nervového systému, kde popisují délku svalu nebo míru změny jeho délky.
Svalové vřeténko se skládá z intrafusálních vláken a extrafusálních vláken. Intrafusální vlákna jsou uložena v pouzdře z pojivové tkáně. Ve svalovém vřeténku se nacházejí dva typy intrafusálních vláken: „nuclear bag1 a bag2 fibers“ a „nuclear chain fibers“.
„Nuclear bag fibers“ jsou schopny rychlé adaptace na vzniklé změny v délce svalu, změny rychlosti a zrychlení prováděné kontrakce. Okolo nich se obtáčí primární aferentní vlákna Ia (rychlá vlákna).
„Nuclear chain fibers“ jsou schopný jen pomalé adaptace. Součástí jejich aferentního systému jsou Ia vlákna, ale i sekundární (keříčkovitá) II pomalá vlákna.
Polární konce svalového vřeténka jsou kontraktilní a jsou drážděny malými gama-motorickými nervovými vlákny, které leží v předních rozích míšních (gama-motoneurony představují 30 % z celkového počtu motoneuronů). Svalové vřeténko je inervováno senzoricky, motoricky a autonomně.
3. Mezi metody, které úspěšně využívají facilitaci svalových vřetének můžeme zařadit postizometrickou relaxaci, metodu excentrické dekontrakce (ED) dle Brüggerova konceptu, metodu sestry Kenny a jiné.
6 SOUHRN
Svaly a jejich tonus jsou kontrolovány pomocí proprioreceptorů (svalové
vřeténko a Golgiho svalové tělísko). Svalové vřeténko vykonává funkci
komparátoru, který stále porovnává vlastní délku s délkou okolních
extrafusálních svalových vláken. Jestliže se sval při kontrakci stahuje, klesá
dráždivost svalových vřetének. Přes gama-smyčku je přitom zajištěno současné
přiměřené zkrácení intrafusální vláken a tím i zachování dráždivosti svalových
vřetének při nové výchozí délce svalu. Kontrakce svalu může být vyvolána buď
přímým podnětem z alfa-motoneuronu, nebo nepřímo – reflexně,
prostřednictvím podnětů z gama-motoneuronu.
Tento mechanismus je využíván při mnoha rehabilitačních
metodách, zejména pak při excentrické dekontrakci (ED) Brüggerova konceptu,
metodě postizometrické relaxace (PIR), metodě dle sestry Kenny.
7 SUMMARY
Muscles and their tone are controlled via proprioreceptors (muscle spindle and Golgi tendon organ). Muscle spindle operates as a comparator, comparing its length to length of neighbouring extrafusal fibres. Excitability of muscle spindles is decreasing during muscle contraction. The maintenance of excitability of muscle spindles at new initial muscle length together with simultaneous and adequate contraction of intrafusal fibres is ensured by gamma-loop. Muscle contraction can be evoked by either direct stimulus from alfa-motoneuron or indirectly by reflex stimuli from gamma-motoneuron.
This
mechanism is used by many rehabilitative methods, especially by excentric
decontraction (ED) of Brügger conception, postizometric relaxation (PIR) and
nurse Kenny's method.
8 REFERENČNÍ SEZNAM
Adler, S. D., Beckers, D., & Buck, M. (1993). PNF in practice: an illustrated guide. Berlin: Springer.
Atwood, H. L., & MacKay, W. A. (1989). Essentials of Neurophysiologiy. Toronto, Philadelphia: B. C. Decker
Inc., 306 – 313.
Bakker, D. A. & Richmond F.
J. R. (1982). Muscle Spindle Complexes in
Muscles Around Upper
Cervical Vertebrae in the Cat. Journal of Neurophysiology. USA: Department of Physiology, Queen´s
University, Kingston, July 1, 62 –
76.
Boyd, I.,
& Gladden, M. (1998). Muscle Spindle
and Strech Reflex. University of Glasgow. Retrieved 21.10.2002 from the
Wolrld Wide Web: http://www.umds.ac.uk/physiology/mcal/spin1.htm.
Božíková, Š. (1999). Možnosti
facilitace u terapie měkkých tkání se zaměření na postizometrickou relaxaci
(PIR). Bakalářská práce, Univerzita Palackého, Fakulta tělesné kultury,
Olomouc.
Brügger, A. (1986). Die
Funktionskrankheiten des Bewegungsapparates. Stutgart: Gustav Fischer
Verlag.
Capko, J. (1998). Základy
fyziatrické léčby. Praha: Grada Publishing.
Holubová, D. (2002). Srovnání efektu dvou technik PNF „Contract –
Relax“ a „Hold - Relax“ u dysfunkčních ischiokrurálních svalů. Diplomová
práce, Univerzita Palackého, Fakulta tělesné kultury, Olomouc.
Deursen, R. W. M., Sanchez,
M. M., Ulbrecht, J. S., &
Cavanagh, P. R. (1996). A Quantitative Test of Muscle
Spindle Function in Diabetic Neuropathy. American Society of Biomechanics. USA, Georgia, Atlanta: Presented at the 20th
Annual Meeting of the American Society of biomechanics, October, 17 – 19.
Enoka, R. M. (1994). Neuromechanical
Basis of Kinesiology. Champaign: Human Kinetics.
Frömel, K., & Vaverka F. (1998). Publikační manuál fakulty tělesné kultury. Olomouc: Univerzita
Palackého v Olomouci.
Guyton & Hall. (2000). Textbook
of Medical Physiology. Philadelphia:W. B. Saunders Company.
Havlíčková, L. (1999). Význam excentrické kontrakce pro posturu. Rehabilitace a fyzikální lékařství, č. 1, 9 – 14.
Hulliger, M. (1983). The
Mammalian Muscle Spindle
and Its Central
Control. Zürich: Brain Research
Institute, University od Zürich.
Hunt, C. C. (1990).
Mammalian Muscle Spindle: Peripheral Mechanisms. St. Physiological Reviews . USA: the
American Physiological Society, July 3,
643 – 663.
Chaitow, L., & DeLany, J. W. (1996). Modern neuromuscular techniques. Edinburgh: Churchill Livingstone.
Janda, V. (1993). Základy kliniky
funkčních (neparetických) hybných poruch. Brno: IDVPVZ, 14 – 15.
Kusunuse, R. S. (1993) Strain and
counterstrain. In J. V. Basmajian, R. Nyberg, Rational manual therapies
(pp. 327 – 329). Baltimore: Williams & Wilkins.
Lewit, K. (1996). Manipulační
léčba v myoskeletální medicíně. Heidelberg-Leipzig: J. A. Verlag,
Hüthig GmbH.
Mayer, M. (1997). Některé
neurofyziologické aspekty spasticity. Rehabilitace a fyzikální lékařství, č. 2. 41 – 46.
Mayer, M. & Konečný, P. (1998). Možnosti ovlivnění spasticity
prostředky fyzikální terapie u rehabilitace nemocných s centrálními
poruchami hybnosti. Rehabilitácia,
31, 40 – 46.
Matthews, M. (2000). Muscle spindle.
Retrieved 5.1.2003 from the Wolrld Wide Web: http://www.sfu.ca/~tmiluer/spinal.pdf.
Opavský, J. (2002). Neurologie. Ústní
sdělení.
Pavlů, D. (1996). Neurofyziologické koncepty v pohybové terapii a
jejich kontroverze. Rehabilitace a
fyzikální lékařství, č. 4, 179 – 182.
Pfeiffer, J. a kol. (1976). Facilitační
metody v léčebné rehabilitaci. Praha: Avicenum: Zdravotnické
nakladatelství.
Rodová, D., Mayer, M., & Janura, M. (2001). Současné možnosti využití
povrchové elektromyografie. Rehabilitace
a fyzikální lékařství, č. 4. 173 – 177.
Seidel, Z., Obenberg, J., & Süssová, J. (2002). Myorelaxancia v ambulanci internisty očima neurologa. Retrieved 29.9.2002 from the Wolrld Wide Web:
http://www.zdrava-rodina.cz/med/med889/med889-31.htm/.
Smékal, D. (1998). Syndrom
bolestivého ramene – diagnostika a kinezioterapie. Diplomová práce,
Univerzita Palackého, Fakulta tělesné kultury, Olomouc.
Trojan, S. a kol. (1996). Lékařská
fyziologie. 2. vyd. Praha: Grada Publishing.
Trojan, S., Druga, R., Pfeiffer, J., & Votava, J. (2001). Fyziologie a léčebná rehabilitace motoriky
člověka. Praha: Grada Publishing.
Vodičková, M. (1996). Porovnání
účinků metod postizometrické relaxace svalové a excentrické dekontrakce pro
ovlivnění zkrácení ischiokrurálního svalstva [Diplomová práce]. Univerzita
Palackého, Fakulta tělesné kultury, Olomouc.